
ヒトが細菌に感染したときには、防御機能として「体液性免疫」が主に働き、「IgG抗体の産生⇒好中球による細菌の貪食」といった流れで細菌を排除します。
しかし、このほかにも細菌をやっつける協力な助っ人が存在しており、それが「補体」という物質なのです。
目次
補体とは
補体は、英語の”complement”の頭文字をとって「C」と表記されます。大まかにC1~C9の9つの成分があり、C1はさらにC1q、C1r、C1sの3つに、その他の補体はa.bの2つの成分などに分けられます。
補体系は20以上のタンパク質とタンパク質断片からなり、主に肝臓で合成され、血中に放出されます。補体は易熱性(熱に弱い)であり、56℃、30分の処理で失活します(=非働化)。
細菌の侵入などをきっかけにして、連鎖的に反応することでさまざまな免疫反応を引き起こします。
抗体が体内に侵入してきた細菌などに結合すると、補体は抗体により活性化され(古典経路)、膜侵襲複合体を形成して細菌の細胞膜を壊すなどして生体防御に働きます。
補体の働きは?
http://www.emeneki.com/
それぞれの補体がなんらかの働きを発揮するためには、その補体がある”きっかけ”によって連鎖的に活性化されることが必要です。
基本的には、C1から順々に活性化していきますが、なぜかC4だけ2番目にくると覚えましょう!
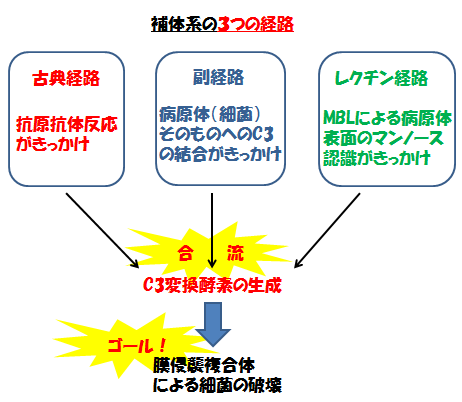
補体系の経路には大きく3つの経路があり、それぞれスタートとなるきっかけが違いますが、途中からは合流するため目指すゴールは同じです。
各段階で活性化した補体には、オプソニン化、好中球の遊走、ウイルス中和などさまざまな機能があります。
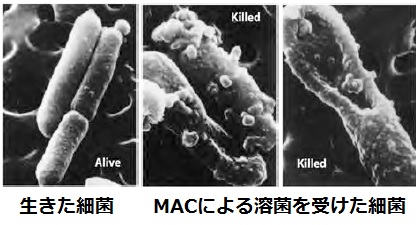
補体系の最終目標は、C5b6789(C5b-9)、別名: 細胞膜障害性複合体または膜侵襲複合体(membrane-attack complex : MAC)を作り上げることです。
この膜侵襲複合体は、細胞膜に穴をあけて壊して細菌を殺す(免疫溶菌現象)という強い作用があり、細菌防御において好中球に負けないくらいの力を発揮します。
【膜侵襲複合体(MAC)の働き】
①古典経路
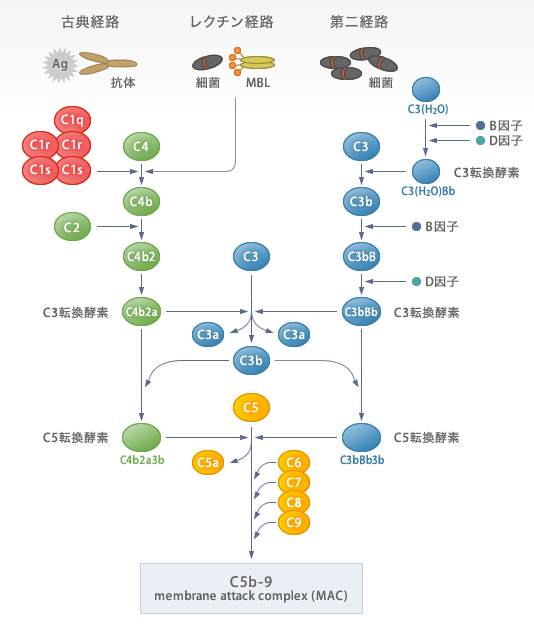
古典経路は、C1からスタートする”the・正攻法”の経路です。古典経路のきっかけは、体液性免疫の抗原抗体反応です。
細菌などの抗原に抗体が結合し、さらに補体(C1)がその抗体にくっつくことで活性化します。
3種類あるC1のうち、抗体に結合する補体はC1qであり、C1qが結合するとC1rが活性化してC1sを分解・活性化します。
活性化したC1sはC4とC2に作用して、それぞれC4aとC4b、C2aとC2bに分解します。C4bは細胞膜に結合できる性質があり、C2aは細胞膜に結合したC4bと結合して複合体(C4b2a)をつくります。
C4b2aは、C3を分解するC3転換酵素として働き、C3をC3aとC3bに分解します。そして、C3bはC4b2aと結合し、C4b2a3bとなってC5を分解して活性化していきます。
というように、ある活性化した補体が他の補体を分解・活性化させ、その一部とどんどん結合していきます。
最終的に、C5~C9が順番に活性化して結合していくとC5b6789(膜侵襲複合体)が形成され、細菌に穴をあけて溶菌します。
②副経路
C3は肝臓で作られて血液中に放出された後、一部が自然に加水分解されてC3(H20)に変化しています。
ここで新しい2種類のタンパク(因子)が登場します。B因子とD因子とよばれているものです。
C3(H20)にB因子が結合すると(C3(H20)B)、さらにD因子により、そのB因子がBaとBbに分解されC3(H20)Bbとなります。
C3(H20)BbはC3変換酵素として働き、C3をC3aとC3bに分解します。つまり、血中では常に一定のC3が分解されてC3aとC3bが生成されているわけですね。
重要なのはC3bのほうですが、これは細菌などに出会わなければすぐに不活性化されてしまいます。
しかし、ちょうど近くに細菌がいたときには細菌表面にくっつき、さらにB因子がC3bに結合します。
ここでもD因子が活躍して、C3bに結合したB因子を分解してC3bBbとなり、これまたC3変換酵素として働きます。
すると、細胞表面ではC3変換酵素の活性がすごく高まってC3⇒C3a+C3bの分解がどんどん進んでいくことになります。
一方、C3bBbは増幅されたC3bとさらに結合してC3bBb3bとなって、C5変換酵素として働き、やっと古典経路と合流します!
その先は、古典経路と同じように補体の活性化が進んでいきます。
③レクチン経路
レクチン経路は、スタートの部分だけが古典経路と違いますが、ほとんど古典経路と同じです。
細菌の表面にはマンナンという成分が存在しています。マンナンとはD-マンノースからなる多糖類です。
レクチン経路は、マンノース結合レクチン (Mannose-Binding Lectin:MBL) 経路ともよばれており、細菌表面のマンノース残基にレクチン(MBL)が結合することによってはじまります。
すると、MBL関連セリンタンパク質分解酵素であるMASP-1とMASP-2(それぞれC1rとC1sに似ている)を活性化し、C4とC2に作用して、それぞれC4aとC4b、C2aとC2bに分解します。
C4bとC2aは古典経路と同じように、互いに結合してC3転換酵素を形成します。古典経路をと比べるわかりますが、「C1」の役割が「MBLとMASP」に変わっただけですね!
覚えておくべき補体!
経路の各段階で生成した補体成分には、それ自体にも免疫作用をもつものがあります。そのなかでも、特に重要な働きをもつ補体をご紹介します。
C3a、C5a
マスト細胞を刺激してケミカルメディエーターを遊離させ、即時型反応(アナフィラキシー)を起こします。(アナフィラトキシンとして働く)
C3b
異物に結合し好中球やマクロファージの貪食能をアップさせます。(オプソニン化)
C5a
好中球を炎症部位に呼び寄せるケモカイン(遊走因子)として働きます。
C5b6789
細菌防御において最も重要な複合体であり、免疫溶菌反応を引き起こします。別名:膜侵襲複合体(MAC)とよばれ、細菌の細胞膜を破壊することで、細菌に穴をあけて壊します。
まとめ
補体は、連鎖的に活性化する20種以上のタンパクの総称であり、細菌防御において「好中球による貪食」とともに重要な役割を果たす。
補体系の活性化には、「古典経路」「副経路」「レクチン経路」の3つの経路が存在している。
それぞれの経路の「活性化のきっかけ」は異なるが、途中で合流し最終的なゴールは「膜侵襲複合体(MAC)」の形成である。
活性化した各段階の補体には、アナフィラトキシン、オプソニン化、ケモカインなど免疫システムで重要な役割をもつものがある。